Anatomia mózgu
Mózg człowieka to ok. 86 miliardów neuronów zorganizowanych w wyspecjalizowane obszary.
Model 3D
Skan T1 — strukturalny rezonans magnetyczny
Rezonans magnetyczny T1 (T1-weighted MRI) to podstawowa technika obrazowania struktury mózgu. Kontrast T1 opiera się na czasie relaksacji podłużnej tkanek: istota biała (mielina) wypada jasno, istota szara — szaro, płyn mózgowo-rdzeniowy — ciemno. Poniższy skan pochodzi z mojego własnego mózgu — wykonano go podczas testu pilotażowego eksperymentu; rozdzielczość izotropowa pozwala rekonstruować dowolne przekroje w płaszczyznach osiowej, wieńcowej i strzałkowej.
Badania fMRI opierają się na tym samym sprzęcie co T1 — różni je tylko sekwencja impulsów: T1 mierzy anatomię, fMRI (sekwencja EPI) wykrywa zmiany przepływu krwi związane z aktywnością neuronów. Skan strukturalny T1 służy jako mapa anatomiczna, na którą nakładane są wyniki funkcjonalne.
Płaty mózgowe
Kora mózgowa (łac. cortex cerebri) dzieli się na cztery pary płatów, odpowiadające czterem kościom czaszki. Każda półkula — lewa i prawa — zawiera wszystkie cztery płaty. Podziały anatomiczne wyznaczają bruzda środkowa (sulcus centralis) oddzielająca płat czołowy od ciemieniowego, bruzda boczna Sylwiusza oddzielająca płat skroniowy, oraz bruzda ciemieniowo-potyliczna.
Płat czołowy (lobus frontalis) zajmuje ok. 1/3 objętości kory i jest proporcjonalnie największy u człowieka. Odpowiada za funkcje wykonawcze, kontrolę impulsów, planowanie i pamięć roboczą. Jego tylna część zawiera korę ruchową pierwotną.
Płat ciemieniowy (lobus parietalis) przetwarza informacje sensoryczne i integruje je z informacją przestrzenną. Zakręt zaśrodkowy to pierwszorzędowa kora somatosensoryczna. Uszkodzenia mogą powodować zaniedbywanie jednostronne — pacjent ignoruje połowę przestrzeni i własnego ciała.
Płat skroniowy (lobus temporalis) odpowiada za percepcję słuchową, rozumienie mowy i pamięć długotrwałą. Hipokamp, ukryty w głębi płata skroniowego, jest niezbędny do tworzenia nowych wspomnień deklaratywnych. Uszkodzenie obustronne hipokampa (przypadek H.M.) powoduje ciężką amnezję.
Płat potyliczny (lobus occipitalis) jest w całości poświęcony przetwarzaniu wzrokowemu. Kora prążkowana (V1) to pierwsze korowe piętro wzrokowe — neurony reagują na orientację krawędzi i kontrast. Informacja wzrokowa biegnie dalej dwiema drogami: grzbietową (gdzie/jak) do płata ciemieniowego i brzuszną (co) do dolnej kory skroniowej.
Kora ruchowa i somatosensoryczna
Po obu stronach bruzdy środkowej leżą dwa sprzężone obszary: kora ruchowa pierwotna (M1) z przodu oraz kora somatosensoryczna (S1) z tyłu. Oba obszary mają somatotopową organizację — każdy punkt na powierzchni ciała jest reprezentowany w określonym miejscu kory.
Homunkulus Penfielda to wizualizacja tej mapy: zniekształcona figura człowieka, w której rozmiar poszczególnych części odpowiada powierzchni korowej poświęconej ich unerweniu. Twarz, usta i dłonie zajmują nieproporcjonalnie duże obszary — ich mięśnie wymagają najdrobniejszej kontroli (mowa, manipulacja przedmiotami). Nogi i tułów mają relatywnie małą reprezentację.
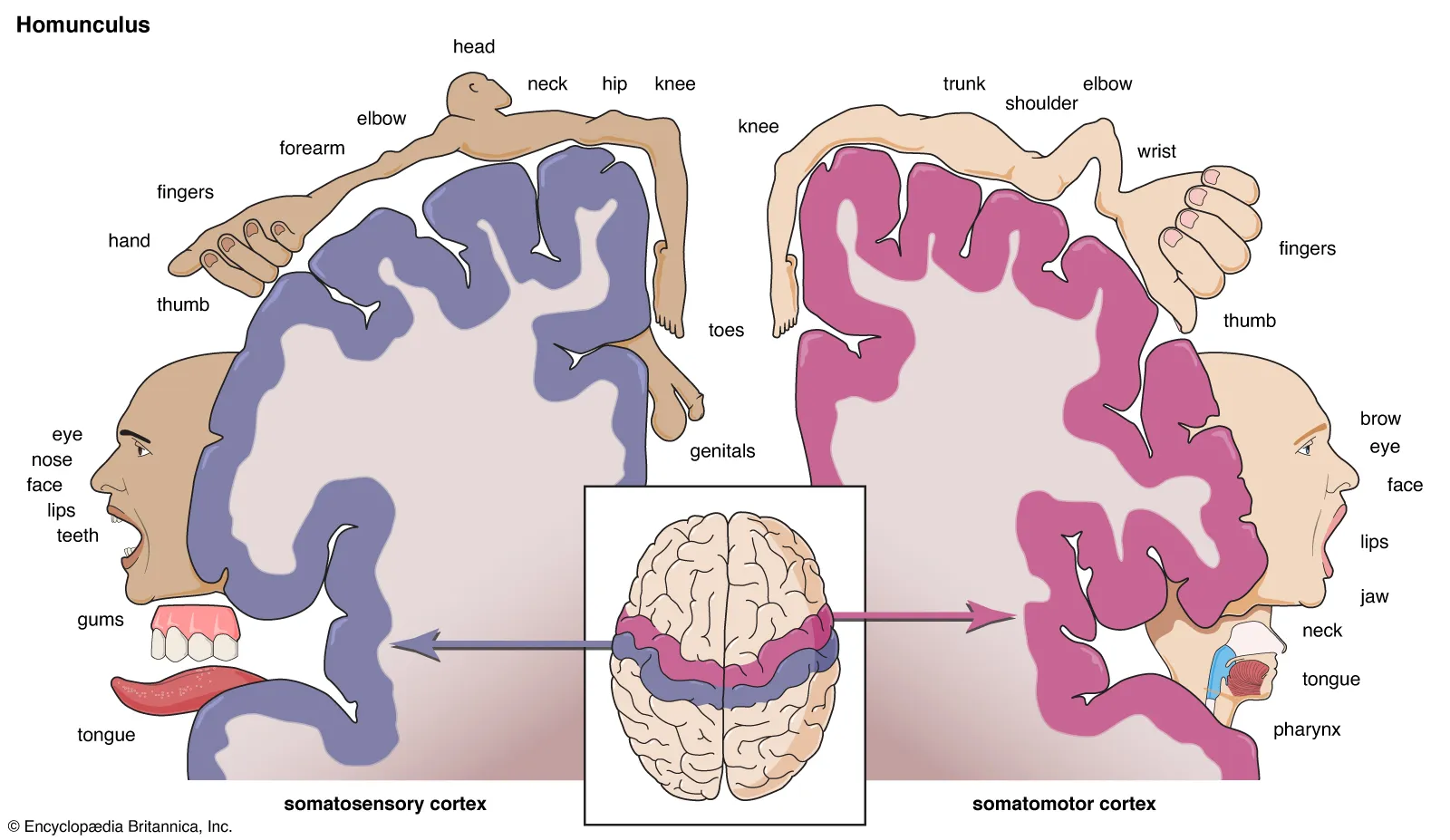
Autorstwo: Encyclopædia Britannica, Inc./Steven N. Kapusta
Źródło: Encyclopædia Britannica (dostęp: 4 czerwca 2026 r.)
M1 zawiera neurony piramidowe Betza — jedne z największych neuronów układu nerwowego. Aksony tych komórek tworzą drogę korowo-rdzeniową (piramidową), skrzyżowaną na granicy rdzenia przedłużonego. Dlatego uszkodzenie lewej M1 powoduje porażenie prawej strony ciała i odwrotnie.
S1 odbiera sygnały z receptorów skórnych, mięśniowych i stawowych za pośrednictwem wzgórza. Neurony S1 są wrażliwe na lokalizację, intensywność i modalność bodźca (dotyk, temperatura, ból). Neuroplastyczność korowa oznacza, że mapa może ulegać reorganizacji — np. u muzyków obszar reprezentujący palce jest powiększony.
Obszary językowe
Język jest funkcją silnie zlateralizowaną — u ok. 95% osób praworęcznych i ok. 70% leworęcznych dominuje lewa półkula. Dwa kluczowe obszary, odkryte w XIX w., pozostają fundamentem neuropsychologii języka.
Pole Broki (Brodmann 44 i 45) leży w dolnej części zakrętu czołowego lewej półkuli. Odpowiada za planowanie sekwencji motorycznych mowy, płynność artykulacji oraz przetwarzanie gramatyczne i składniowe. Uszkodzenie powoduje afazję Broki: chory rozumie, ale mówi z wysiłkiem — mowa jest telegraficzna, pozbawiona spójników i końcówek fleksyjnych.
Pole Wernickego (Brodmann 22) leży w tylnej części górnego zakrętu skroniowego. Odpowiada za rozumienie mowy i dobór właściwych słów. Uszkodzenie daje afazję Wernickego: chory mówi płynnie i z właściwą intonacją, lecz mowa jest pozbawiona sensu (liczne parafazje, neologizmy, „sałatka słowna") i nie rozumie wypowiedzi innych.
Oba obszary połączone są pęczkiem łukowatym (fasciculus arcuatus). Jego uszkodzenie powoduje afazję przewodzenia: rozumienie i spontaniczna mowa są stosunkowo zachowane, lecz chory nie potrafi powtórzyć usłyszanego zdania.
Współczesne badania fMRI i stymulacja przezczaszkowa pokazują, że sieć językowa jest szersza — obejmuje też korę przedruchową, wyspę, zakręt kątowy i zakręt nadbrzeżny. Model Broki–Wernickego jest uproszczeniem, ale użytecznym punktem wyjścia.
Kluczowe eksperymenty
| Cel | Lokalizacja funkcji mowy w mózgu. |
| Procedura | Badanie pośmiertne mózgów pacjentów z afazją — niemożnością płynnej mowy przy zachowanym rozumieniu. Kluczowy przypadek: Leborgne (zwany „Tan"), który przez 21 lat wypowiadał jedynie sylabę „tan". |
| Wyniki | Uszkodzenie w tylnej części dolnego zakrętu czołowego lewej półkuli korelowało z afazją niefluowną u wszystkich badanych pacjentów. |
| Wnioski | Mowa jest zlokalizowana w lewej półkuli. Był to pierwszy empiryczny dowód na lateralizację funkcji i modularność mózgu — przełom w neuropsychologii. |
| Cel | Mapowanie somatotopowej organizacji kory ruchowej i somatosensorycznej przed neurochirurgią padaczki. |
| Procedura | Elektryczna stymulacja kory mózgowej u przytomnych pacjentów podczas operacji. Pacjenci opisywali doznania lub wykonywali ruchy przy podrażnieniu poszczególnych punktów kory. |
| Wyniki | Systematyczna mapa ciała na korze — homunkulus ruchowy i somatosensoryczny. Nieproporcjonalnie duże obszary dla twarzy i dłoni, odpowiadające gęstości unerwienia ruchowego i czuciowego. |
| Wnioski | Kora ruchowa i somatosensoryczna mają somatotopową organizację. Wyniki opublikowane w książce „The Cerebral Cortex of Man" stały się podstawą neurochirurgii funkcjonalnej. |
| Cel | Zrozumienie roli hipokampa w tworzeniu wspomnień długotrwałych. |
| Procedura | Przypadek Henry'ego Molaisona (H.M.) — pacjenta z obustronnymi resekcjami hipokampa w leczeniu ciężkiej padaczki. Milner przez dziesięciolecia prowadziła szczegółowe testy neuropsychologiczne H.M. |
| Wyniki | H.M. stracił zdolność tworzenia nowych wspomnień deklaratywnych (amnezja anterogradalna) przy zachowanej pamięci proceduralnej i pamięci z przed operacji. Inteligencja i osobowość bez zmian. |
| Wnioski | Hipokamp jest niezbędny do konsolidacji pamięci deklaratywnej (epizodycznej i semantycznej). Pamięć proceduralna jest niezależna od hipokampa. Rozróżnienie systemów pamięci stało się fundamentem neuropsychologii. |
Pamięć i hipokamp — przypadek pacjenta H.M. · YouTube
| Cel | Badanie funkcji każdej półkuli po przecięciu ciała modzelowatego. |
| Procedura | Pacjenci po komisurotomii (przecięcie spoidła wielkiego w leczeniu padaczki) — mózg „split-brain". Prezentacja bodźców do jednego pola widzenia (tj. jednej półkuli) i pomiar reakcji ruchowych każdej ręki oddzielnie. |
| Wyniki | Lewa półkula przetwarza mowę i przetwarzanie werbalno-analityczne; prawa dominuje w przetwarzaniu przestrzennym i całościowym. Każda półkula działa jak niezależny mózg — brak wzajemnej świadomości po przecięciu połączeń. |
| Wnioski | Dwie półkule mózgu mają wyspecjalizowane, komplementarne funkcje. Ciało modzelowate jest niezbędne do ich integracji. Sperry otrzymał Nagrodę Nobla w 1981 r. |
Split-brain: badania nad przecięciem ciała modzelowatego · YouTube
Pacjent H.M.
Henry Molaison (1926–2008), znany w literaturze jako H.M., to najczęściej opisywany pacjent w historii neuropsychologii. W 1953 roku William Beecher Scoville wyciął mu obustronnie hipokampy w leczeniu ciężkiej padaczki. Operacja była skuteczna. Skutki uboczne okazały się trwałe.
H.M. przestał tworzyć nowe wspomnienia deklaratywne. Każde spotkanie z Brendą Milner, która badała go przez dziesięciolecia, wyglądało dla niego jak pierwsze. Pamięć proceduralna pozostała nienaruszona: uczył się nowych umiejętności motorycznych, choć nie pamiętał, że się uczył.
Jego przypadek udowodnił, że pamięć nie jest jednorodna. Hipokamp konsoliduje fakty i zdarzenia, ale nie ruchy ani nawyki. Praca Milner z H.M. rozdzieliła te dwa systemy na dobre.
Patient H.M. and the discovery of memory systems · YouTube
Ślepota nieuwagowa
Ślepota nieuwagowa (inattentional blindness) to zjawisko, w którym osoba nie zauważa wyraźnego bodźca w polu widzenia, bo jej uwaga jest skierowana na inne zadanie. Najbardziej znany przykład to eksperyment Simonsa i Chabrisa (1999) — „niewidzialny goryl". Uczestnicy liczyli podania piłki i aż połowa z nich nie zauważyła osoby w stroju goryla przechodzącej przez scenę.
Zjawisko to dobitnie pokazuje, że percepcja nie jest biernym rejestrowaniem — widzenie wymaga uwagi. Poniższe filmy to klasyczne demonstracje tego efektu i jego wariantów.
Test selektywnej uwagi — niewidzialny goryl · YouTube
Test uwagi — moonwalking bear · YouTube
Kto dokonal morderstwa? Eksperyment z uwagą · YouTube